- 中新世における狭鼻猿の共存関係
- 中務真人(京都大学・理・自然人類)
- 中新世の狭鼻猿類のコミュニティーを概観し、霊長類の共存関係というテーマについて考察する。タフォノミーに関わる問題は大きく、断定的な議論は難しいが、以下のような点が指摘できる。
- 中新世前期の東アフリカは温暖であった。ケニアを中心とする各化石サイトからは同所的に数種類の化石真猿種が発見される(図1、表1)。サイズの異なる複数の狭鼻猿種によってコミュニティーが形成されていたと考えられる。豊かな資源に加え、サイズの変異、食性の違いが、複数種の類人猿の共存を可能にしたのではないか。一方、中新世中期には、小型の類人猿が顕著に減少する。また、オナガザル上科が類人猿と一緒に現れるようになる。類人猿とオナガザルの同所性には地域差があるが、サイトあたりの狭鼻猿種数は明らかに減少する。ナチョラでは霊長類化石のうち、類人猿が大半だが、マボコではビクトリアピテクスが大半である。ただし、類人猿の多様性がアフリカ全体で実際に低下したかどうかは、東アフリカ以外で発掘が進まないと不明な点がある。こうした変化には、乾燥化、季節性の亢進が関係していると考えられる。アフリカでは、オナガザル上科は中新世前期中葉までにはあらわれている。しかし、種数が増え始めるのは鮮新世以降である。中新世前期から中期の東アフリカで、類人猿が実際に衰退したとすれば、最大要因は環境の変化だったと考えるのが妥当ではないだろうか。オナガザルとの競合的関係はあったとしても、二次的要因であり、ことさらに種間競争を主張する必要はないと筆者は考える。
- ヨーロッパでは異なる種類の化石狭鼻猿の同所性はほとんど見られない。生息環境の分断化による結果なのか、それとも排他的な関係があったかは、系統関係と拡散のタイミングに不明な点が多く断定できない。しかし、類人猿同士、プリオピテクス同士、類人猿・プリオピテクス、いずれについても同所性が著しく低いこと、その一方で、食性の変異幅は東アフリカ程度に大きかったと推定されることを合わせ考えると、単なる地理的隔離だけでは説明できない。
-
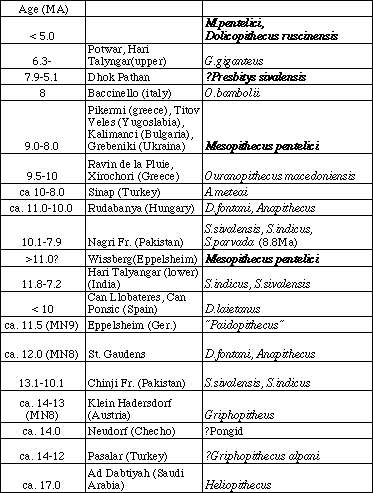
シワリクでは複数のシバピテクスが同所的に棲息している。しかし、はっきり同定できるシバピテクスの種数自体が少ないため、東アフリカとの類似性など、共存の意味については不明な点が多い。シバピテクスの主なものには現在3種知られているが、もっぱら、サイズで区別され、形態的には類似性が高い。仮に種間の生態差が小さかったとすれば、なぜ400万年間、シワリクでは複数種が共存したかという疑問が残る。
- オナガザルのユーラシアへの拡散は、ホミノイドの絶滅が始まる直前に起こっている。また、最初期の出土地域は極めて限定されている。こうしたことから、オナガザルの進化がヨーロッパの類人猿の絶滅に直接の影響を与えたとは考えられない。また、同じ事はシバピテクスの絶滅とギガントピテクスの進化についても言えるであろう。
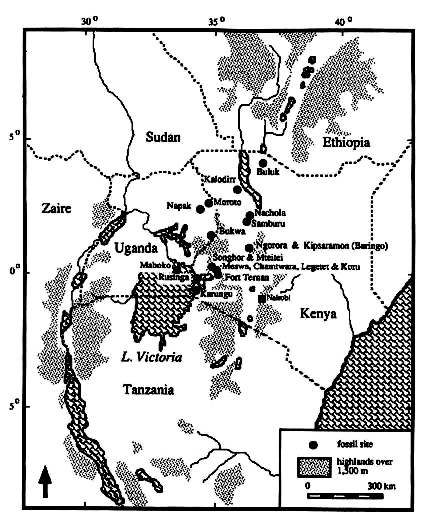
図1 東アフリカにおける中新世霊長類化石サイト。Klein (1989) Human Career. The University of Chicago Press, Chicago, より。
表1 東アフリカの化石狭鼻猿類のサイト、年代、分布(太字はオナガザル上科)

図2 ヨーロッパ、ランドママルエイジ9-10(11.5-9.0MA)における狭鼻猿類の分布。Andrews et al. (1996) Distribution and biochronology of European and southwest Asian Miocene catarrhines. In (Bernor, Fahlbusch and Mittmann eds.) The Evolution of Westren Eurasian Neogene Mammal Faunas Columbia Univ. Press, New York, pp.168-207 より。
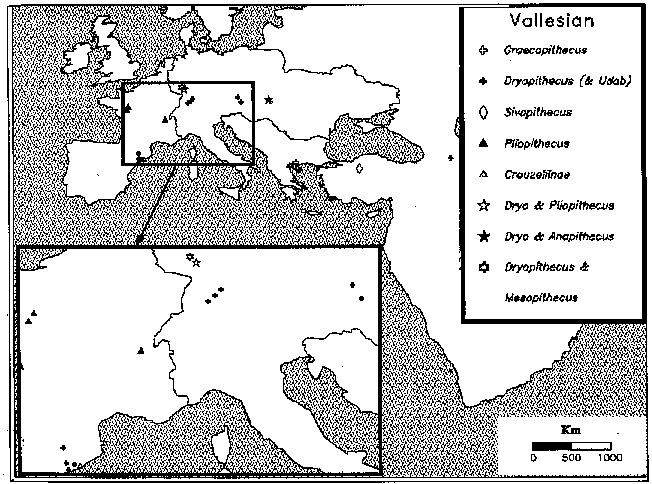
表2 ヨーロッパ・西アジア・シワリクの化石狭鼻猿類(プリオピテクス類は省略)のサイト、年代、分布(太字はオナガザル上科)
公開シンポジウムのページに戻る