- 初期人類における種分化と同所性について
- 諏訪 元(東京大・総合研究博物館)
- 初期人類の系統分類は近年、細分傾向がはやりのようである。例えば、初期のホモ・エレクトスは独自の種エルガスターとされ、種数全体としては初期エレクトスとラミダスを含め12以上がもうけられる(筆者は10種とする)。極端な細分の例では、例えばアファレンシスやエルガスターが複数種に分けられる。また、属レベルでもアルディピテクス、アウストラロピテクス、パラントロプスの他、プレアントロプスが用いられる。
- では、これらの細分傾向は実際の種、属レベルにおける多様性を示しているのだろうか?研究者によっては、ホモ属の出現時に少なくとも3種への分化があったとし、直立ニ足歩行を伴ったヒト科そのものの出現期にも同様な、あるいはそれ以上の系統的放散があったのではと憶測している。化石の記録はそもそも多様性を過小評価するはずだし、種分化が起こっても骨や歯の形態的分化を伴わないことも多かろうとの議論が細分主義の背景にあるようだ。しかし、こうした細分された初期人類の属や種に基づく多様性は、場合によっては「見かけ上」のものであり、生物学的種のレベルにおける多様性とは必ずしも同一視することはできない。
- 属レベルの分類はそもそも人為的であり、ここでは特に議論しない。通常は、近縁性と共に、形態と行動生態からみてそこそこにまとまりのある適応様態をもってして一つの属とする。筆者らは広義のアウストラロピテクス属を用い、パラントロプス属は設けていない。これは、頑丈型猿人もそうでないアウストラロピテクスも、大なり小なり頑丈な咀嚼器で特徴づけできることと、頑丈型猿人が単系統群であると未だに言いきれないことへの配慮からである。
- 種レベルの細分は、一つには種の概念によって左右される。近年、細分を提唱する多くの研究者はphylogenetic species conceptに追従する。この種の概念は、一口で言うと、系統的に分析可能な形態単位をもって種とする。このため、種内の多型は実質上みとめられず、現生種の亜種レベルに相当する下位分類群もが種と認識されることにつながる。
- では、そもそも古生物記録において種をどうとらえるのか?もし、現生種における「生物学的種」と整合する尺度を求めるならば、種内の多型をみとめたうえで過去の多様性を見積もるのがのぞましい。前述の、種分化が起こっても骨や歯の形態的分化を伴わない可能性の問題は、その都度考察すべきことであり、予め細分に傾く理由とはなり得ない。
- 実際、高等霊長類で同所的に生存する近縁種が骨と歯で区別できない実例があるのだろうか。筆者はこれを調査する機会を得ていないが、もしあるとするならばグエノン類の中だけだろう。グエノン類の多くは派手な顔面模様で知られるが、これが種の認識に機能するとも言われている。骨格の差異は少なくして、異同の明瞭なシグナルとなっているのかもしれない。
- 野外調査の進展に伴い、高等霊長類の場合、自然状態で近縁種間に雑種が形成されることが明かにされてきた。高等霊長類のように、種ごとの生殖生理に強い特色もなく、一般に生殖行動も柔軟な場合、むしろ当然なのかもしれない。では、いわゆる種間に交雑がある場合、もはや種でなく亜種とみなすのが妥当だろうか?有名なヒヒの例がさんざん議論されてきたと思う。特に印象的なのは、ゲラダヒヒとアヌビスヒヒが平気で交雑し立派に子孫を残し、しかも、明らかな適応度の低下を一見伴わないことである。集団遺伝学的にみた場合、こうした交雑が「種」間の遺伝子交流につながるのか、それとも実質的にはそうした効果がないのか、私は不勉強で知らない。しかし、現生ゲラダヒヒの行動生態の独自性と、ゲラダ属の400万年近い系譜を考えた場合、表現型レベルでは系統的独自性を十分保ってきたと言ってよいだろう。
- このように生殖的隔離が完璧であることが望めない以上、「種」を考える場合、交雑の有無より系統ごとの独自性が重要であろう。そうなると、例えばマントヒヒとアヌビスヒヒが亜種か種かといった問題も、今後、双方が行動生態、形態などにおいてそれぞれの独自性を継承して行くかどうかにかかっていることになる。もし、ポピュレーション単位で様々な混合に至り、地域集団以上の広がりにおいて表現型レベルの特色が流動的に変貌し、時代ごとに再編されるならば亜種と考えたほうが良いだろう。こうなると、種、亜種の区別は場合によっては進化史的結末でしか定まらないことになる。即ち、種間交雑は、その可能性は常に存在するのだが、それぞれの種のもつ行動生態の指向性故、起こらないのである。種を維持するための「実質的な生殖隔離」は生物側の「主体性」に依存するとでも言えよう。
- そして骨格系をも含めた形態的分化(すなわち風貌、顔貌、体格、体型の変化)がこれと関連する場合もあろう。こうなると、古生物記録は断片的であるために不利、不正確であるどころか、進化史を見極めて判断できるという大きな利点を持つことになる。
- こうした観点から初期人類の系統をみてゆくと幾つかの点が浮かび上がる。ます、同一遺跡から出土する化石の微々たる形態差から主張されている複数種の同時代、同所的生存は甚だ疑わしい。こうした議論はアフリカヌス猿人、アファール猿人などほとんどの猿人の種でみられる。特に注意を要するのは、同一遺跡群の「同時代」の化石群集には、多くの場合1000から10000年、場合によっては100000年オーダーの年代幅が含まれているため、環境変遷やポピュレーション移動を考えると、複数亜種の混合であってもおかしくないことである。
- また、突拍子もない系統論が正しくない限り、初期人類の近縁種(もしくは細分した「種」)間の分岐の深さは、深い場合でも100万年程度にしか見積もれないことに注意したい。おおよそは現生ヒヒの亜種間程度の遺伝的違いと同等であろう。ゴリラとチンパンジーの分岐ははるか700万〜800万年前以上前に遡るとすると、両者の共存は初期人類の同所的生存とはそもそも事態が異なることが明らかである。
-
初期人類においても、生殖行動はそこそこに柔軟であったとすると、同所的に共存する2種はそれなりにはっきりした形態的分化を遂げていたか、あるいは相当はっきりとニッチェの分化をおこしていたはずである。いずれにせよ、300万年前以後の頑丈型猿人とホモ属の系統(もしくはその祖先種)にそれぞれ属する2種が、東アフリカでも南アフリカでも、時代ごとに同所的に共存したのである(図)。それ以外には、複数種の明らかな同所的共存の証拠はないと私は思っている。
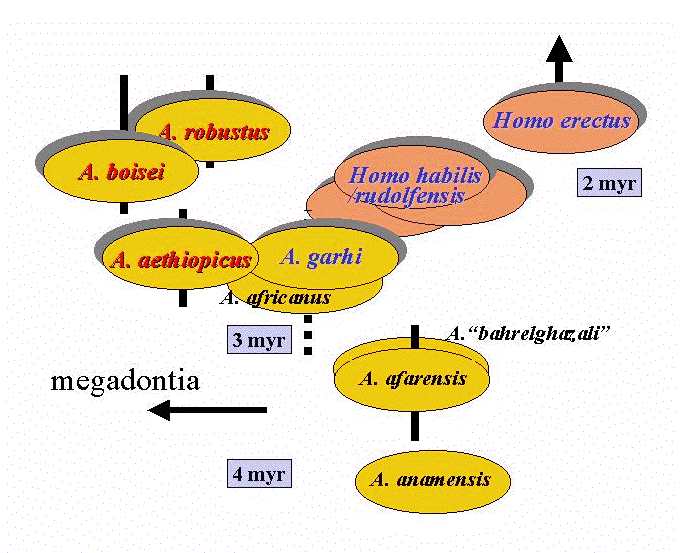
- 最初の2種共存は、東アフリカで約270万年前から250万年前ごろにガルヒ猿人とエチオピクス猿人に見られる。1999年に我々が発表したガルヒ猿人の資料はエチオピアのアファールから出土したもので、そこでは頑丈型猿人は発見されていない。しかし、従来から知られているオモとトゥルカナの断片的な歯の化石群の中にはエチオピクス猿人のものと、大型だが頑丈型でないのものとがあり、おそらくエチオピクスとガルヒの同所的共存を示しているものと思われる。以後、東アフリカでは初期からエレクトスまでのホモ属とエチオピクスもしくはボイセイ猿人の共存があり、南アフリカではホモ・ハビリスもしくはエレクトスとロブストス猿人の共存がある。200万年前を下ると形態的に、そしておさらくニッチェ的にも分化が相当はっきりしてくると言えよう。
- 問題はこの2系統が分化したばかりの時代である。形態的分化、ニッチェ分化はどうだったのか?このころは頑丈型猿人でなくとも、そもそも咀嚼器が頑丈になって行った時代である。食性傾向としては、各猿人の種とも大方似ていたことも十分考えられる。ここで少し古環境の変遷を概観してみたい。海洋底コアの研究などから、300万〜250万年前ごろの間に、高緯度地域では寒冷化を伴いながら南北両極の氷床が激しく増減するようになったと考えられている。そして、アフリカの低緯度地域ではこれが乾燥化として現れ、季節性が増したとも推測されている。こうした環境変動の中、初期人類は種を問わず咀嚼器を頑丈にする傾向にあった可能性がある。そして採食が厳しくなる中、ホモ属の系統では打製石器を多用する方向で適応し、頑丈型猿人の系統ではさらに咀嚼器を頑丈にしていったのだろう。
- しかし、ガルヒとエチオピクス猿人の食性が根本的に異なり、採食活動の重複が全くなかったとも思えない。と言うのは、「頑丈な」咀嚼器の利点とは、機能的には堅い食物の分断、分断効率の向上、磨耗への耐性などにあると考えられるからである。即ち、そもそもはサバンナのモザイクにおける万能型として幾度か独自に生じた可能性が高い。
- もしそうならば、当初の二系統のニッチェ分化は不完全であり、主として季節的に特色を持っていたと考えるのが妥当かもしれない。この場合、種の独自性を保持することが何故可能だったのか?そもそもは異所的に分化したのであろうが、270万年前ごろまでには少なくともトゥルカナでは同所的に生存したらしい。比較的短期の異所的隔離と思われる。エチオピクス猿人に既にみられる、頑丈型猿人独特の顔面形態を伴った形態分化が種の維持と関連したのかもしれない。逆に、頑丈型猿人は早期から極端に特殊化した食性適応を遂げており、ニッチェ分化がそもそも強ったのかもしれない。行動生態と認知科学の分野からの提言を頂きたいところである。形態進化を追及すると共に、こうした多面的な考察により、初期人類の進化史を解きほぐして行きたいものである。
公開シンポジウムのページに戻る