- �R�����g�@�����쒷�ނ̎�ԊW�̌����Ɛl�ސi���_�|����Ɖۑ�|
- ��@���i���s��w�E���E�l�ސi���_�j
- ���͂��߂�
�@�ސl����q�g�̐i���ɂ����āA��ԊW�̉e�����l����Ƃ��̃|�C���g�ɂ��āA�쐶�쒷�ނ̌����T�C�h����A��̃R�����g���������B��ԊW�ɂ́A��q�U�z��َ�Ԃ̃R�~���j�P�[�V�����Ƃ������A��Ԃŗ��v�������炷�悤�ȋ��i�������肤�邪�A�����ł͌����쒷�ނ̎�Ԃ̋����𒆐S�ɍl���Ă݂悤�B�܂��A��̓I�ȍޗ��́A�q�g�Ȃ̐i���Ƌ��ʐ����l���₷���A�A�t���J�̗쒷�ނł̎�ԊW�Ɍ��肵�Ă���B
- �����_�I���_
�@���݂̗쒷�ނ̓������ɂ͂Q�̃��x������ʂł���B�ЂƂ́A���Y�̕����킪����n���ɕ��z���A�嗤���x���̕��z�n�}�Ȃǂł͏d�����ĕ��z���Ă���ɂ�������炸�A�~�N���Ȑ����ꏊ�ł́A���݂킯�Ă��铯�����ł���B���������ꍇ�́A�����I������Ƃ����Ă��A���������ɂȂ邱�Ƃ͂܂��Ȃ��B�����ЂƂ́A�~�N���Ȑ����ꏊ�ł����݂킯�Ă��炸�A�����킪�H����x���ꏊ�ȂǂŁA���ʂ��������𗘗p���āA����I�ɏo��悤�ȓ������ł���B�S�����ƃ`���p���W�[�Ȃǂ̑命���̗쒷�ނ̓������́A��{�I�ɂ͌�҂ŁA�َ퓯�m�������œ����ɍ̐H�����邱�Ƃ�������B�����ŁA�������L���ł������A�������������I�ȕ�����̊Ԃɂ͐��ݓI�ɂ͋���������ƍl������̂ł���B
- �@�߉���́A���ɑ���v�������Ă��邽�߂ɁA��Ԃł̓����I�ȋ����́A�ŏ����̂��݂킯���N����Ɨ��_�I�ɂ͍l������B�܂�A�����Ɍ��肪����ȏ�́A�ʂ̎������������������B���邩�A�������鎑�����������I�ɗ��p�ł��鐫�������B����Ƃ����_���ł���B�܂��A�i���j�I�ɂ��݂킯���ꍇ�ɂ́A�`���u���⋣���r����������Ƃ����B�����Œ��ӂ��ׂ��͈ȉ��̓_�ł���B
- �@���������邩��Ƃ����Ă������Ɍ`���u����ُ����ɂނ����Đi������킯�ł͂Ȃ��B�ʂȎ����𗘗p�ł���A�h�{�������������ɂ��ނ��A�ɐB�ɂ܂ʼne�����Ȃ����������肤��B�Ƃ��ɁA�쒷�ނ̂悤�ɁA�H�ו��◘�p�ꏊ����V�I�Ɋw�K�����ł́A�w�K�ɂ���ĉ���\�ȋ����͑I�����ɂ͂Ȃ肦�Ȃ��B����ɁA�쒷�ނƂ��ɗސl���̏ꍇ�ɂ�₱�����̂́A�o�Y�Ԋu�������Ȃ�悤�ɐi�����Ă������Ƃł���B�܂�A���Ƃ��������h�{�����̈����ɂȂ������Ƃ��Ă��A�o�Y�p�x�̒ቺ�ł��ޒ��x�̋����ł���A�������p��ς����ɋ�������������A�o�Y�p�x���Ⴍ�R�h���̖ʓ|����蒷���݂鐫�������̂��I������邱�Ƃ��l������B�o�Y�p�x�̌����Ƃ����i���X���́A��ԊW�Ɍ��炸�A�Љ���w�I�Ȕ��z�ŗސl����l�ނ̐i�����l�@���邤���ł̊�{�I�Ȗ��_�ł�����B
- �@�܂��A�߉��킪�����I�ɐ������A�������p�ɂ��݂킯���݂��邩��Ƃ����āA���ꂾ���ł͎�Ԃ̋����̂��i���̌��ʂƌ��Ȃ����Ƃ͂ł��Ȃ��B�`���u�������������ƍl�@����ɂ́A�Q��̌`�Ԃ̍����A�ُ��I�ɕ�炵�Ă���n��ł��������I�ɕ�炵�Ă���n��ł̂ق����傫�����Ƃ�������Ȃ��Ă͂Ȃ�Ȃ��B
- �@�`�ԂɌ��炸�A���̗��p�ɂ�����鐫���ł���A�H����s���p�^���ɂ����Ă��A���l�̘_���ŁA���������퍷�̊g���i���I�ɂ����炷�ƍl���Ă����������͂Ȃ��B�������A�쒷�ނł́A����ł�����߂đ��l�Ȋ��ɐ������A�����������̗��p�\�����n��ɂ���đ傫���قȂ邤���ɁA�H���₻�̑��̍s���p�^���ɂ͔��`�I�ȗv�����t�s���Ă���B�H����s���X�������̓����ɑΉ����Ă���Ƃ��Ă��A���ꂪ��`�I�ł��邱�Ƃ������邽�߂̗��_�I�Șg�g�݂͏\���Ƃ͂����Ȃ��B����́A��ԊW�ɂ����炸�A�쒷�ނ́u�Љ�Ԋw�v�̍��{�I�Ȗ��_���B
- �@�������L���ŋ����̉e��������̂��ǂ�������������ɂ́A���ϓ��̕����l���邱�Ƃ��K�v�ł���B�����ʎ��H���Ȃǂ̊��͈��łȂ��A�G�߂�N�ɂ���đ傫�ȕϓ�������B���������āA�쒷�ނ̎�ԊW�ɋ����I�ȑ��ʂ��������Ƃ��Ă��A�����͂������ʂ����킯�ł͂Ȃ��A�����ʂ����Ȃ��Ȃ�G�߂�N�ɖ��ƂȂ�B��ԊW���l���邤���ł́A�����̕ϓ����������A�{�g���l�b�N�ɂȂɂ�������̂����������邱�Ƃ��d�v�ł���i�O�J2000�j�B��ʂɔN�Ԃ�ʂ��Ĉ��肵�Ă���ƍl������M�їтł��A�G�ߕϓ���N�X�ϓ��͂����ŕ�炷�����ɂƂ��đ傫���e������B���Ƃ��A�ʎ��̗ʂ��ނ́A��N�̂����ł��傫���ϓ����A�N�ԂŐ��{�ɂ��Ȃ�G�ߍ�������B�ǂ̒n��̔M�їтł��A�쒷�ނ̂Ƃ��Ă̎����ɋG�ߕϓ������邱�Ƃ͋��ʂ���B�������A�ǂ̋G�߂ɂǂꂭ�炢�������Ȃ�̂��́A�n��ɂ���đ傫���قȂ�A�P���ȍ~���ʂ�C���̔N���ςȂǂɂ���Ă͎����ʂ̗\�����ł��Ȃ����Ƃ��킩���Ă����B��ԊW�̋G�ߕϓ����A�ʎ����Y�ʂȂǂ̎����ʂ�Ή������Č������錤���́A���㔭�W���邾�낤�i����2001�j
- ���T���Ɛl��
�@�l�ސi���_�ɂ́A�������V���ȍ~�̃I�i�K�U����Ȃ̓K�����U�ɂ���āA�����̉e�����������ސl�������ނ��iAndrews 1981�j�A����ɒn�㐫�̃I�i�K�U���Ƃ̋������A�T�o���i�ɐi�o�����ސl���̃q�g���𑣐i�����Ƃ����������i���c1986�j�B�������V���ȍ~�̉��Ώ��^�ސl���̎퐔�̌������A���̉����̊�Ղł��邪�A�����_�ł͉������x�����鉻�̏؋��͎ア�i����2001�j�B���̉����̑Ó������A�����쒷�ނ̎�ԊW���猟�����邱�Ƃ��K�v�ł���B
- �@�ߋ��̃I�i�K�U���Ƒ�^�ސl���ŋ������z�肳���̂́A�ǂ���̌�������ʎ����d�v�ȐH���Ƃ��Ă��邩��ł���B�ސl���͑�^�Œn�㐫�������A����A�I�i�K�U�����Ȃ́A��^�������ɁA���Q����������A�j�܂����B���A�ʎ��̐H�Ɍ����I�Ȑ����������A�ސl���Ƃ͈قȂ���ꉻ���݂���i�ۋ�1996�j�B�܂�A�o�����ʎ����߂��鋣���ɗR������`���u���������\������������̂ł���B
- �@�ʎ��H�ł̋������`���u���������炵���Ƃ����l�����������鎖���ɂ́A�R���u�X���ȂƃS�����̗t�H�ւ̓��ꉻ������B�ǂ���̎���A���@���ނ̂Ȃ��ł͗t�H�̌X���������A�R���u�X�ł͎��̌`������ǂɁA�S�����ł͏����ǂ⒰���ۂɓ��ꉻ���݂��Ă��āA���l�̐i���v���Z�X����������B�R���u�X�ƃS�����́A���̕��z�⓯���I�ȃT���̎�ނ�\���ɂ͑Ή��W������̂�������Ȃ����A�����͂���Ă��Ȃ��B�R���u�X�ɂƂ��Ă͎���̗t���ς��A�S�����ɂƂ��Ă͒n�㐫�̑��{���d�v�ȐH���ł���B����ƒn��ɂ��݂킯���邩����A�R���u�X�ƃS�����͋������\�ł���̂�������Ȃ��B�����ŁA�S�����ɂƂ��ďd�v�ȐH�������ł���͂��̒n�㐫���{���قƂ�ǂȂ��A�K�{���̑吼�m�݂̔M�їт̏��������Ă݂悤�B���̒n��̃S�����́A���n��̃S�����ƈ���ĂقƂ�ǂ̃l�X�g������ɂ���A�n�㐫���{�̂����ɁA�t���ς���v�ȐH���Ƃ��Ă���炵���A���n��̃S���������㐫�������\���������B���������āA�R���u�X�Ƃ̐��ݓI�ȋ����������Ɨ\�z�������ł���B���ہA���̕ی��ɂ́A�I�i�K�U���͂S�킢��ɂ�������炸�A�R���u�X���܂��������Ȃ��̂ł���B�S�����ƃR���u�X�ɋ��ʂ���@�ې��H���Ƌ����I�ȑ��ʂ́A���コ��Ɍ��������ׂ���ԊW�ł��낤�B
- �@�`���p���W�[�̓��H�X���̐i���ɂ��A�T���ނƂ̋����̉e����z��ł���B�`���p���W�[�́A�Ƃ��ɃT�����悭�����̂ŁA���H�ɂ̓T���Ƃ̋����ڌ��炷���ʂ�����B�Ȃ��ł��A�J�R���u�X�́A�`���p���W�[�̑�D���ł���A�R���u�X�̌̐��̓`���p���W�[�̎�ɂ���ė}�����Ă���B�܂��A�A�J�R���u�X�̍��Q�`���́A�`���p���W�[�̐ڋ߂����m���₷�����A�ߐH�������헪�ł���Ƃ�������x������؋������A�t���J�̃`���p���W�[�ł͓����Ă���(No� & Bshary 1997)�B�������A���̂悤�ȍ��Q�̋@�\�́A���A�t���J�ł͊m�F�ł����A�i���j�I�ȌX���ƍl����ɂ́A�܂��܂������̗]�n������i�ܕS��2000�j�B
- �@�����ȊO�ɂ��A�ސl���̃T���Ƃ̋����͂����߂�悤�ȍs����̓����́A�����������Ƒz�肳���n��Ŕ��B���Ă��邩�ǂ������A�����ސl���̒n��Ԃł̔�r���猟������K�v������B
- �������q�g�Ȃ̋���
�@�����q�g�Ȃ̕�����Ԃł̋����ɂ�������̍����́A���^�Ɖؚ��^�̓�̃^�C�v�̓������ł���i�z�K2001�j�B���^�Ɖؚ��^�̓����I�ȋ����́A���A�t���J�k�������A�t���J�̍L���͈͂̊e���łQ�T�O���N�ȏ�O����P�O�O���N�O��ɂ�����܂Ő������Ă����Ƃ����B�����̃q�g�Q�^�C�v�̋����́A���^�̐�łɏI����Ă���B���̂��Ƃ���A���̂Q��Ԃł͋������s�\�Ȃ����́A�����������������ƍl�������Ȃ邪�A���ۂɁA���������ǂ̃��x���̎��Ԃ��ԂŖ��ł������̂��́A�܂��悭�킩��Ȃ��B��q�̂悤�ɁA���Ƃ��Ɠ����n��ɕ�炵�Ă��Ă��A����I�ɂ͈قȂ�A���^�C�v�𗘗p���Ă����̂ł���A�����͎����I�ɂ͖��ɂȂ�Ȃ��B
- �@���̖����l����ɂ������āA���f���Ƃ��ׂ��M���́A�������q�g�ɋ߉��Ȍ����̃S�����ƃ`���p���W�[�̊W�ł���B�`���p���W�[�ؚ͉��^�ƁA�S�����͊��^�Ƌ��ʓ_������A�����l�ނ̋�������������̂ɂ����Ă��̍ޗ��ɂȂ�B
- �@�����̃q�g�Ȃ̂Q�^�C�v�ƌ����ސl���̊Ԃőz��ł��鋤�ʓ_�́A��^�Œn�㐫�������A�A���H�ւ̈ˑ����������Ƃł���B�`���p���W�[�͓��H���A������g�p���ĉh�{������ς��邱�Ƃ��ł���_�ł������q�g�ȂƂ̋��ʐ��������A�`�ԓI�ɂؚ͉��^�Ɏ��Ă���B����A�S�����͒n�㐫���`���p���W�[���������A���̓_�ł̓`���p���W�[���������q�g�ȂɎ��Ă��邾�낤�B����ɁA�S�����́A�͂����͂ő̊i�����傫���_�����^�ƌ`�ԓI�ɋ��ʂ���B�ސl���ԂƏ����q�g�ȊԂ̃p�������Ȑi���X���́A�e��̓����ɁA�\���I�Ɉ��肵���g�ݍ��킹������Ƃ�������i���I�Ȍ��ۂ�z�肳����B
- ���S�����ƃ`���p���W�[�̋���
�@�����̃S�����ƃ`���p���W�[�̕��z�͍L���d�����Ă���i�}�P�j�B���A�t���J�ł́A�R���S���勤�a���̓����ŁA�܂��A�����A�t���J�ł́A�i�C�W�F���A�A�J�����[���A�K�{���A�ԓ��M�j�A�A�����A�t���J���a���A�R���S���a���A�A���S���k���̍L���͈͂ŋ������Ă���B���A�t���J�ɂ̓`���p���W�[�������������A������͂Ȃ����A�قƂ�ǂ̃S�����́A����̋�����ɂ���̂ł���B
- �@�S�����ƃ`���p���W�[�̋����ƁA�I�i�K�U���̕�����ł̋����Ƃ̋��ʓ_�Ƃ��ẮA�@�H���̏d�����傫���A�����ɉʎ��Ń��j���[���d������A�A��ԓI�ɂ��݂킯�Ȃ����A�ʎ��������R�����G�߂ł����݂ɔr�����Ȃ��A�B�����͏��Ȃ��A��{�I�ɂ͌��G���Ȃ��A�Ƃ������_���w�E�ł���B
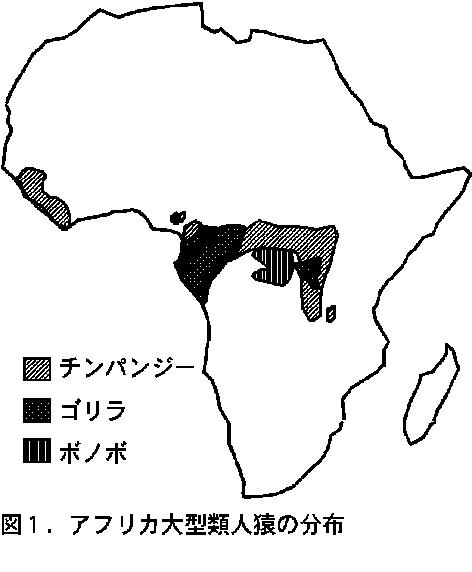
- �@���������M�ђn��̗쒷�ނ́A�����I�ɐ���ނ��������邱�Ƃ����ʂł���A�P�Ƃł��邱�Ƃ͋H�ł���i���c1999�j�B�������������̓����́A�A�t���J�ސl����I�i�K�U���ނɌ��炸�A�A�W�A��A�V���E�U���ł������ނˋ��ʂ��Ă���B�܂�A�쒷�ނ́A����Ȃ�W�c�ԂŔr���I�ɂȂ�悤�Ȏ�ł����Ă��A�܂��A�߉��ł����ɑ���v�������Ă��Ă��A�َ�Ƃł���Ί�{�I�ɂ͋����\�ł���B���������āA���Η쒷�ނł��A�����I�ȗ쒷�ނ̋߉���ɁA�S���d���̂Ȃ��H����s�������Ă͂߂���A�`�ԓI�ɂ͐H���̏d�����K�R�ł������Ƃ��Ă��A��������r���I�ȋ������l����K�v�͕K�������Ȃ��B
- �@����A�T���̋����W�ɑ��ėސl���̎�ԊW�̓����Ƃ��ẮA�@���Q������Ȃ��A�A����Ƃ���^�ŃT�������L���V������g���A�n�㐫��������������n��܂ŁA�����̋�Ԃ��T�����m�����L���A�B�ސl���̋����͂Q��ԂɌ����A�T�����m�ł̂悤�ɑ���ނɂȂ�Ȃ��A�C�S�����ƃ`���p���W�[�̐������x�͕����L��������P���O��ƒႭ�A�T���̂悤�ɕ����L�������萔�\���̖��x�ŕ�炷���Ƃ͂��肦�Ȃ��A�Ȃǂ���������B���������āA�����͈͂ƌ���ꂽ��Ԃł�������̎킪�N���ꏏ�ɕ�炷�T�����m�����A�ސl�����m�͋�����������邽�߂̕����I�ȗ]�T���傫���ƍl������B���Ȃ킿�A���̈����ɂƂ��Ȃ��āA�������������Ȃ��Ă��A�ސl���Ԃł̓T���Ԃ�����Ԃ̋����̈��e����������₷���̂�������Ȃ��B
- �@���ΐl�ނł��A�����ސl���Ɠ��l�ɁA�����I�ȋ߉��퐔�͏��Ȃ��A�������x���T����肸���ƒႩ�����ƍl�����Ă���iKelley, 1994�j�B�����ŁA�����ސl�����m�̋������T�����m�̋����ƈقȂ�_������ɖ��炩�ɂ��Ă������Ƃɂ���āA�����l�ނ̋����W�̓����������ƍl�@�\�ɂȂ邾�낤�B
- �@�S�����ƃ`���p���W�[�̋�����͔M�їтł���B�M�їтł̐������x�́A��Ԃ̋����̂��߂ɁA�S���������̒n���`���p���W�[�����̒n��̐������x�����Ⴂ�ƍl�����Ă������Ƃ�����B�������A�P���ɍl����A���̊��ɑ���v���͎��Ă���̂ŁA�L���Ȓn��ł́A����Ƃ��������āA�`���p���W�[�ƃS�����̐������x�͑��ւ���Ɨ\�z�����B�Ƃ͂����A���ꂼ��̎�̕��z�̒[�����ł́A�`���p���W�[�����̊��ƁA�S���������̊������邱�Ƃɂ܂������͂Ȃ��A���ۂɈ���̎�݂̂���������n�������̂ŁA�P���ɗ���̖��x�����ւ��邱�Ƃ͂��肦�Ȃ��B�S�������`���p���W�[�����\�N����������Ă��邱�Ƃ��l����ƈӊO�Ȃ��Ƃ����A����̖��x�̑��ւɂ��ẮA�܂��������ł���B�ߔN�A�����̋����n��ŁA���ʂ������@�ŗސl���̖��x���肪�s���Ă��Ă���A�߂������ɖȖ��Ȍ������\�ɂȂ�B
- �@����ɔM�їтƂ����Ă��A�n��ɂ���đ��l�ł���A���F���قȂ�B�ސl���͉ʎ��ɑ���n�D���������A�܂��A�n�㐫�̑��{���d�v�ȐH���ɂ��Ă��邪�A�ʎ����Y��n�㐫�̑��{�̗ʂ́A�e�n�̔M�їтő傫���قȂ��Ă���B�Ɠ����ɁA�S�����ƃ`���p���W�[�̐������x���e�n�̔M�їтňقȂ�̂����A�ʎ���n�㐫���{�̂ǂ������������A�������x�Ɍ����Ă���̂��܂��킩���Ă��Ȃ��B����ɁA�ǂ̋�����̒����n�ł��A�S�����ƃ`���p���W�[�̐H���́A�ʎ����p�𒆐S�Ƃ��đ傫���d�����A���p�ꏊ�����݂���B�����̉\���͍����͂��ł��邪�A�ʎ��̗ʂ���ނ����Ȃ������������ɂ̓S�������@�ې��H���ɋ����ˑ�����悤�ɐH�����V�t�g���邱�Ƃɂ���āA���������͋N�����Ă��Ȃ����Ƃ��������Ă���(Kuroda et al, 1996)�B
- �@�S�����ƃ`���p���W�[�Ԃł̋����ɐi���I�ɈӖ������������Ƃ������邽�߂ɂ́A�P���ɗ���̐����ɂ��݂킯�I�ȑ��Ⴊ�F�߂��邾���ł͕s�\���ł���B������ł̂��ꂼ��̗ސl���̐H����s���������A�P�Ƃŕ�炵�Ă���n��ł̐��������퍷��傫������悤�ɃV�t�g���Ă���ǂ�������������K�v������B�H���ɂ��Ă͌����\�ɂȂ���邪�i�R�z2000�j�A������̗ސl���̍s�������ɂ��Ă͂܂��s���ȓ_�������A����̌����̐i�W�����҂����B�܂��A�O�q�̂悤�ɁA�ސl���̐������͑��l�ł���A�H����s���ɂ͔��`�I�Ȑ����������̂ŁA���̍��ɑΉ������ސl���̐����̍����A�ǂ��܂Ői���I�ȈӖ����������������邽�߂̗��_�I�Ȑi�W���K�v�ł���B
- �@�����q�g�Ȃ̋������ސl���̋����ƈقȂ�_�́A�ސl���͔M�їт𗣂�Ă͓����I�ɂ͋����ł��Ȃ����A�q�g�̑c��͊����n�тŋ����\�������ƍl�����Ă���_���B�����̃q�g�Ȃ��ސl���ƈقȂ�ƍl�����鐫���́A���s�Ƃ����ɒ[�Ȍ`�ԓI�ȓ��ꉻ�A����ɂƂ��Ȃ������n�㐫�ƁA�����n�ł̐����ł���B�M�їт𗣂ꂽ�ꍇ�̋����́A�����ސl���ŕs�\�ł���悤�ɁA�����l�ނɂƂ��Ă��ނ������������낤�B�������A���ۂɂ͋������B������Ă����̂ł���A���̂��Ƃ͋�����ۏ����Ղ��A�ސl���Ƃ͑傫���قȂ��Ă������Ƃ��Ӗ����Ă���B���s�ƒn�㐫���A������e�Ղɂ��闝���͂����ɂ͎v�����Ȃ��B�����n�ł́A�A�����̎�v���������Ȃ����߁A�����l�ނ̎�Ԃł́A�`�ԓI�Ȏ퍷�������̗ސl�����x�ł���Ƃ���Ȃ�A�ސl���̎�Ԃł̑���ȏ�ɐH�����قȂ��Ă������A�܂��́A�����ސl���ɂ��܂��Ė��x���Ⴉ�������A����������邽�߂̋��͂ȎЉ�I�������������ɈႢ�Ȃ��B���������l�@��i�߂邽�߂ɂ́A�S�����ƃ`���p���W�[�̋�����̕Ӊ����ł̎�ԊW���A����ɒNj������ׂ��ł���B
- ���ސl���ƃq�g
�@�S�����ƃ`���p���W�[�̑c��́A�����I�ɕ�炷���ƂɂȂ�����A�����̃q�g�ȂƂ����ݓI�ɂ͋�������ł��낤�B�����̗ސl���ƃq�g�́A�ǂ�����A�����H���ɋ����ˑ����Ă���A�q�g�ł��A�X�т�����A�X�тŎ��̏W������B�����̃q�g�Ȃ̋����̑���́A�X�тɈˑ����悤�Ƃ����Ƃ��ɂ����ɂ����ސl���ł������̂�������Ȃ��̂��B�������A����������ԊW�́A����܂łقƂ�ǒ��ڂ���Ă��Ȃ�(Tutin et al., 1996)�B�����̗ސl���Ԃł̊W�Ɠ����ɁA��̏W���Ɨސl���̊Ԃł́A�H�����Ώۂ̏d���Ⓖ�ړI�Ȍ��p�^���Ȃǂ���������K�v�����邾�낤�B
- ���T���^���f��
�@�����l�ނ̊����n�ł̎�Ԃ̋����ɂ��Ẵ��f���ɂ́A�T�o���i�����L�[�ƃp�^�X�����L�[�̑g�ݍ��킹������B���̂Q��́A�S�����ƃ`���p���W�[�̕��z�Ƃ͂قƂ�Ǐd�Ȃ炸�A�Z�l�K������G�`�I�s�A�A�^���U�j�A�܂ł̍L���͈͂ɏd�����z����B���z��̌��݂̐A���̓T�o���i�Ƒa�J�тŁA�l�ސi�����i�Ƃ����悤�Ȋ����n�ł���B����Ƀ��f���Ƃ��Ă̖��͂ɂ́A�p�^�X�����L�[�̌`�ԓI�ȓ����ɂ�����B���Ȃ킿�A�ړ��\�͂������p�^�X�����L�[�̒����㎈�́A�����n�ō̐H���������߂�K���ł���A�������A�z���G���N�g�X�ł̌㎈�̔��B�Ƀp�������ł���ƍl���邱�Ƃ��ł���(Isbell et al., 1999)�B�܂��A�`�Ԃ���ꉻ�������p�^�X�����L�[�́A���̈ړ��\�͂ɂ������H���ƍ̐H�p�^���ɓ��ꉻ���Ă���i����2000�j�B����̃T�o���i�����L�[�̓p�^�X�����L�[�Ǝ����H���������A�p�^�X�����L�[���������ŁA�V����͋����A�I�i�K�U���Ƃ��Ă̌`�Ԃ�H���ɓ��ꉻ�݂͂��Ȃ��B
- �@�ǂ���̎���I�i�K�U�����Ȃ̓����ł����n�I�ȏW�c�����邱�Ƃ͋��ʂ��邪�A�p�^�X�����L�[�͐��I��^���傫���P�Y�Q������̂ɑ��āA�T�o���i�����L�[�͐��I��^�����������Y�Q������B����A�ǂ���̎���o�Y�Ԋu���Z���A���������ɑ��Y�Ƒ��n�œK��������ł���Ƃ���Ă��āA�o�Y�����グ�邱�Ƃ�����ȑ�^�̃T����ސl���̓K���Ƃ̑Δ䂪�����[���B
- �@����ɁA�����l�ނ̉������@����Ă������A�t���J�Ɠ�A�t���J�́A�p�^�X�����L�[�̕��z�Ƃ��܂�d�Ȃ��Ă��Ȃ��B����T�o���i�����L�[�́A�`�ԓI�ɂ̓p�^�X�����L�[�̂悤�ȓ��ꉻ���ア���A���z�̓p�^�X�����L�[�Ƃ͈قȂ�A�����l�ނ̉��ΎY�n�̓��A�t���J���A�t���J�ɂ��L�����z����B�܂��A�T�o���i�q�q�́A�T�o���i�����L�[�Ɠ��l�ɁA�����n�ւ̌`�ԓI�ȓ��ꉻ���キ�A��n�I���Y�Q������A�T�o���i�����L�[�Ɠ����悤�ɃT�n���ȓ�̊����n�тɍL�����z����B
- �@�����̃T���̕��z��`�Ԃ�Љ�I�����ƂƂ��ɁA��ԊW�ɂ́A�����l�ނ̐i���ƃp�������Ȍ��ۂ������Ɗ܂�ł���ɈႢ�Ȃ��A�ސl���݂̂Ȃ炸�A�T���̓������ɂ��Ă��A����Ɍ������K�v�ł��낤�B
- ���܂Ƃ�
�@����܂ŁA�����̗쒷�ނ̌����́A�킲�Ƃɍs���邱�Ƃ������������߁A��Ԃ̊W�ɏœ_�����Ă������͂܂��܂����ꂩ�甭�W�̗]�n������B���������A�쒷�ނ̐������x���e�킲�Ƃɒn��Ԃłǂ̂悤�ɑ�������̂��A�܂��A�e��̊Ԃłǂ̂悤�ɑ��ւ���̂����A�قƂ�ǂ킩���Ă��Ȃ��B�����P�O�N�̊ԂɁA�e�n�̗쒷�ނ̃o�C�I�}�X�̒l�����肳��A����������ԊW�̉ۑ�́A�����\�ɂȂ����iFleagle et al. 2000�j�B
- �@�ސl���Ə����l�ނ̎핪�����ǂ��ł����ǂ̂悤�ɂ��ċN�������̂��́A���̎�������͈ˑR�Ƃ��Ė��𖾂ł���B�Ƃ͂����A�ߔN�̂c�m�`�ɂ��n���W�̎������ςݏd�Ȃ��Ă��āA�q�g�Ɨސl���Ԃ݂̂Ȃ炸�A�S�����̎�i����j�̕����A�`���p���W�[�̎�i����j�̕����̎������l�@�\�ɂȂ��Ă����B���Ώ؋�����̏��������ɂӂ������ʁA�ސl���̕����ƁA�q�g�̊��^�̏����A�ؚ��^�̔]�̊g��A�����x�̑��i�A�Ί�̏o���Ȃǂ��A�Q�O�O���N��O�サ�āA���ƏW�����Ă������Ƃ�����������B���̎����ɃA�t���J�S�̂ɑ傫�ȋC��ϓ����������ƍl�������Ȃ邪�A���̕ϓ��ɂ���Ċe�킲�Ƃɐi�������Ƃ݂Ȃ��̂ł͂Ȃ��āA��ԊW���ӂ��߂āA�l�ސi����ސl���̎�i����j�����Ƃ����Č������邱�Ƃ��K�v�ł���B
- �@�����쒷�ނ̎�ԊW�́A���ꎩ�̂����ł��ʔ������A��ԊW�̐i���j�I�ȃp�^�����������邱�ƂŁA�����l�ނ̕�����̋����ɂ��Ă͂���ɗL�]�Ȏ����������邾�낤�B
- �����p����
�����O2001�D�쒷�ނ̍��Q�ɂ������ԊW�D�i���l�ފw���ȉ�j���[�X���^�[No. 2: 25-28.
Andrews P, 1981. Species diversity and diet in monkeys and apes during the Miocene. In Stringer C (ed.), Aspects of human evolution, Taylor & Francis, London, pp. 25-61.
Fleagle JF, Janson C & Reed KE, 2000. Primate Communities, Cambridge University Press, Cambridge, pp. 329.
�ܕS���T2000�D�A�J�R���u�X�`���p���W�[�|�쒷�ނɂ�����H���|�H����̊W�D���R�K�ەҒ��C�w�쒷�ސ��Ԋw�x�Cpp. 61-84.
Isbell LA, Pruetz JD, Lewis M & Young TP, 1998. Locomotor activity differences between sympatric patas monkeys (Erythrocebus patas) and vervet monkeys (Cercopithecus aethiops): Implications for the evolution of long hindlimb length in Homo. Am. J. Phys. Anthropol., 105: 199-207.
Kelley J, 1994. A biological hypothesis of ape species density. In Thierry B, Anderson JR, Roeder JJ, & Herrenschmidt N (eds.), Current Primatology: Vol. I: Ecology and Evolution, pp. 11-18.
���c���� 1998�D��ԊW�D���c����E�㌴�d�j�Ғ��C�w�쒷�ފw���w�Ԑl�̂��߂Ɂx�Cpp.114-137.
Kuroda S, Suzuki S, Nishihara T, & Oko RA, 1996. Sympatric chimpanzees and gorillas in the Ndoki Forest, Congo. In: McGrew WC, Marchant LF & Nishida T (eds.), Great Ape Societies, Cambridge University Press, pp. 71-81.
�ۋ����1996�D�M�їтɂ�����쒷�ނƉʎ��̋��i���D�Ȋw�C66�F853-861�D
�O�J�돃2000�D�M�їтƗ쒷�ތQ�W�D���R�K�ەҒ��C�w�쒷�ސ��Ԋw�x�Cpp. 15-36.
���쏮�j2000�D�H���̎퍷�Ɛ����|�p�^�X�U���ƃ^���^���X�U���D���R�K�ەҒ��C�w�쒷�ސ��Ԋw�x�Cpp. 177-199.
�����^�l2001�D���V���ɂ����鋷�@���̋����W�D�i���l�ފw���ȉ�j���[�X���^�[No. 2: 28-30.
���c���K1986�D��Z�v���|�V���ƒ�Z�̐l�ގj�D�V�j�ЁD
No� R & Bshary R, 1997. The formation of red colobus-diana monkey associations under predation pressure from chimpanzees. Proc. R. Soc. Lond. B, 264: 253-259.
�z�K��2001�D�����l�ނɂ�����핪���Ɠ������ɂ��āD�i���l�ފw���ȉ�j���[�X���^�[No. 2: 30-33.
Tutin, CEG & Oslisly R, 1995. Homo, Pan and Gorilla: co-existence over 60 000 years at Lop� in central Gabon. J. Human Evolution, 28: 597-602.
�R�z��2000�D�A�t���J�ސl���̃\�V�I�G�R���W�[�D���R�K�ەҒ��C�w�쒷�ސ��Ԋw�x�Cpp. 109-127.
���J�V���|�W�E���̃y�[�W�ɖ߂�